
Пошарова топографія. Шкіра тильної поверхні пальців тонка, підошвової — щільна, особливо в ділянці проксимальної фаланги. Підшкірна жирова клітковина на тильній поверхні пальців розвинена слабко, на підошвовій пронизана сполучнотканинними перетинками та має виражену комірчасту будову. Тильний апоневроз пальців укріплений сухожилками м'язів-розгиначів які кріпляться до фаланг пальців. З підошвового боку сухожилки м'... Читати далі...
15:20 Патологія ядра |
Ядро клітини (nucleus) — це система генетичної детермінації та регуляції білкового синтезу.
Усі клітини організму людини мають ядро. Винятком є високо-спеціалізовані клітини крові — еритроцити, які втрачають ядро в процесі свого розвитку Клітини мають одне (більша частина клітин організму) або два ядра (клітини печінки), зустрічаються і багатоядерні клітини (остеокласта). Форма ядер, як правило, сферична або еліпсоїдна. Однак є ядра, які мають паличкоподібну, бобовидну, сегментовану або неправильну форму. В кожній клітині об'єм ядра та цитоплазми перебувають у певних співвідношеннях, що має значення при оцінці її функціонального стану. При поліплоїдії (кратному збільшенні набору хромосом) об'єм ядра збільшується. Структура і хімічний склад ядра в клітині, що не ділиться (інтерфазній), та під час її поділу (мітозу) мають свої особливості. Основними компонентами інтерфазного ядра є нуклеолема, нуклеоплазма, хроматин, ядерце. Нуклеолема (nucleolemma), або ядерна оболонка, складається із зовнішньої і внутрішньої ядерної мембран. Вони розділені перинуклеарним простором, ширина якого дорівнює 25 нм. Нуклеолема несуцільна, бо має в собі пори (поровий комплекс), через які здійснюється взаємодія між ядром та цитоплазмою. Пори виконують також структурно-організаційну роль, бо з їх допомогою фіксуються інтерфазні хромосоми. При патологічних станах кількість порових комплексів змінюється. Найпоширеніші пошкодження ядерної оболонки пов'язані з розширенням перинуклеарної цистерни, збільшення об'єму якої супроводжується аналогічними змінами канальців едоплазматичної сітки, що, очевидно, пов'язано з циркуляцією внутрішньоклітинної рідини і процесами клітинного набряку. Як правило, розширення перинуклеарного простору є локальними і виникають в результаті гіпоксії. Вони можуть досягати великих розмірів, наприклад, в ендотеліоцитах кровоносних капілярів міокарду при вродженних вадах серця — до 1000 нм. У цих випадках може спостерігатися розрив зовнішньої мембрани. При іонізуючому опроміненні ядерна оболонка частково або повністю руйнується. Нуклеоплазма (nucleoplasma) представлена колоїдним розчином білків, який створює середовище для швидкої дифузії метаболітів, а також для переміщення інформаційної і транспортної РНК. При дії патологічних факторів у нуклеоплазмі виявляються електроннопрозорі ділянки з агрегованими в них скупченнями гранул РНП (рибонуклеопротеїдів), а також набухання ядра. При цьому спостерігаються зміни колоїдно-осмотичного стану ядра. Хроматин (chromatinum) — це комплекс ДНК і асоційованих з нею білків. На ультраструктурному рівні він має вигляд хромосомних фібрил товщиною 20-25 нм. Під час поділу клітини хроматин інтенсивно фарбується, що пояснюється його конденсацією, тобто утворенням тісно скручених (спіралізованих) низок, які називаються хромосомами. В інтерфазному ядрі такий вигляд хроматину отримав назву гетерохроматину. Він зосереджений, головним чином, на периферії ядра біля внутрішьої мембрани ядерної оболонки — периферійний хроматин. Інтерфазне ядро людини містить 46 d-хромосом або 92 s-хромосоми. Кожна d-хромосома складається з двох s-xpoмосом (після удвоєння ДНК в S-періоді). Еухроматином, або деконденсованим хроматином, називаються ділянки хромосом, які скла-даються з деконденсованої ДНК. Якщо гетерохроматин в інтерфазному ядрі є генетично неактивним, то еухроматин визначає процеси траскрипції, тобто активно спрямовує синтез білка. Конденсація еухроматинових структур відображає інактивацію активних ділянок транскрипції і розглядається як патологічне явище, передвісник загибелі клітини. При вираженому набряку клітини спостерігається дискомплексація хроматину, який у вигляді великих грудок накопичується біля ядерної оболонки. Такий феномен у клітині має назву маргінація хроматину (мал. 9).  Ядерце (nucleolus) — це щільна структура округлої форми, в якій відбувається синтез рибосомної РНК (рРНК). В ядрі може бути одно або кілька ядерець. В ядерці є особливо компактна, з фібрилярною консистенцією, зона (фібрилярний центр). Такі ділянки називаються ядерцевими організаторами і саме в них міститься велика кількість копій генів, які кодують рРНК. Існування ядерцевого організатора в кількох хромосомах дозволяє пояснити наявність в окремих випадках в одному ядрі більше одного ядерця.
Розміри ядерця відображають ступінь його функціональної активності. При високому рівні синтезу рРНК розміри ядерця збільшуються. Поблизу ядерця виявляється своєрідне тільце, яке називається тільцем Барра, або статевим хроматином. Було встановлено, що тільце Барра відповідає одній із двох Х-хромосом клітин осіб жіночої статі. Інша Х-хромосома деконденсована і переносить спадкову інформацію.
Збільшення розмірів та кількості ядерець свідчить про підвищення їхньої функціональної активності. Ядерця, в яких переважають гранули над фібрилярною субстанцією, є відображенням різного функціонального стану як ядерець, так і клітини в цілому. Такі ядерця зустрічаються в молодих плазматичних клітинах, активних фібробластах, гепатоцитах, у багатьох клітинах пухлин. Дисоціація ядерець відображає їхню гіпогрануляцію і може бути результатом гальмування ядерцевої транскрипції.
Дезорганізація (сегрегація) ядерець свідчить, як правило, про повне і швидке припинення ядерцевої транскрипції: ядро зменшується в розмірах, спостерігається виражена конденсація ядерцевого хроматину, відбувається розподіл гранул та протеїнових ниток. Ці зміни зустрічаються при енергетичному дефіциті в клітині.
Зміна форми ядра є суттєвою діагностичною ознакою: деформація ядер при дистрофічних процесах та запаленні (грануломатоз), поліморфізм їх при пухлинному рості (клітинний атипізм). Форма ядра може змінюватися також у зв'язку з утворенням численних інвагінацій ядра, що зумовлює збільшення його поверхні і свідчить про підвищену синтетичну активність.
Поряд зі змінами форми ядра при патологічних станах може змінюватися і їхня кількість. Багатоядерні клітини виникають різними способами. По-перше, при злитті клітин, наприклад, велетенські багатоядерні клітини чужорідних тіл та клітини Пирогова-Лангханса, які утворюються при злитті епітеліоїдних клітин.
По-друге, при розладі мітозу відбувається поділ ядра без поділу цитоплазми, що спостерігається після опромінення, при введенні цитостатиків, злоякісному рості. Зустрічаються також клітини, в яких біля незміненого ядра розташовуються маленькі подібні до ядра структури, так звані "супутники ядра" (каріомери, маленькі ядра). Причиною їхньої появи вважають хромосомні мутації. Такі структури виявляють у клітинах злоякісних пухлин при наявності великої кількості фігур патологічних мітозів.
Включення ядра поділяються на справжні і вірусозумовлені. Перші містяться в каріоплазмі і складаються з речовин, що зустрічаються в цитоплазмі (білок, глікоген, ліпіди та ін.). У більшості випадків вони проникають із цитоплазми в ядро через непошкоджені або пошкоджені пори ядерної оболонки чи через пошкоджену ядерну оболонку. Можливе також проникнення цих речовин при мітозі. Приклад — включення глікогену в ядрах печінки при цукровому діабеті ("ядерний глікоген", "дірчасті пусті ядра"). Вірусозумовлені ядерні включення (так звані тільця ядерних включень) нагадують кристалічну сітку віруса і включають в себе білкові часточки, які виникають при внутрішньоядерному розмноженні віруса. їх розглядають також як прояв реакції пошкодження вірусом цитоплазми ("реактивні включення"). Мітоз (mitosis), або каріокінез, непрямий поділ — універсальний спосіб поділу клітин. Він забезпечує неперервність існування клітинних популяций та організмів, точний розподіл генетичного матеріалу між дочірніми клітинами, створює умови для спеціалізації клітин (диференціюючий мітоз), детермінує збільшення кількості клітин у процесі регенерації тканин. Цей процес умовно ділять на 4 фази: профаза, метафаза, анафаза,телофаза. Профаза. Головна подія профази — утворення веретена поділу клітини. Метафаза. У метафазі закінчується утворення веретена поділу, процес конденсаціі хромосом продовжується. Під час метафази хромосоми з'являються в екваторіальній площині, утворюючи метафазну пластинку. Анафаза. На початку анафази хромосоми переміщуються від екваторіальної площини до полюсів клітини. Телофаза — це поділ клітини на дві дочірні (цитокінез і цитотомія). Зупинка мітозу можлива при переході від С2-періоду до початку мітозу. В цей час у клітині може відбуватися наступна реплікація ДНК. Це призводить до збільшення хромосомних наборів у 4-8 або в більше число разів. Морфологічно такі ядра нічим не відрізняються від диплоїдних, але мають більший об'єм. Зупинка мітозу може відбуватися також у профазі або метафазі. Нарешті, клітина може пройти всі стадії мітозу, але не зазнати цитотомії. В останньому випадку виникають клітини з двома ядрами (гепатоцити). Внаслідок ендорепродукції утворюються велетенські поліплоїдні клітини, наприклад, клітини червоного кісткового мозку — мегакаріоцити. Вважають, що поліплоїдні клітини можуть виникати при злитті (остеокласта). Функціональне значення поліплоїдії полягає в забезпеченні збільшення активності клітин у життєвому циклі організму після народження. Розрізняють такі типи патології мітозу (Алов І.О.,1972).
I. Пошкодження хромосом: 1) затримка клітин у профазі; 2) порушення спіралізації та деспіралізації хромосом; 3) фрагментація хромосом; 4) утворення мостиків між хромосомами в анафазі; 5) раннє роз'єднання сестринських хроматид; 6) пошкодження кінетохора. II. Пошкодження мітотичного апарату:
1) затримка мітозу на стадії метафази; 2) розосереждення хромосом в метафазі; 3) тригрупова метафаза; 4) порожниста метафаза; 5) багатополюсеві мітози; 6) асиметричні мітози; 7) моноцентричні мітози; 8) К-мітози. III. Розлад цитотомії: 1) передчасна цитотомія; 2) затримка цитотомії; 3) відсутність цитотомії. Патологія мітозу може виникати під впливом різних патогенних агентів: ультрафіолетове та іонізуюче опромінення, висока температура, хімічні речовини, у тому числі канцерогени і мітотичні токсини та ін. Велика кількість патологічних мітозів спостерігається при малігнізації тканин.
|
|
|
 |
Розділ: | Категорія: Топографічна анатомія нижньої кінцівки
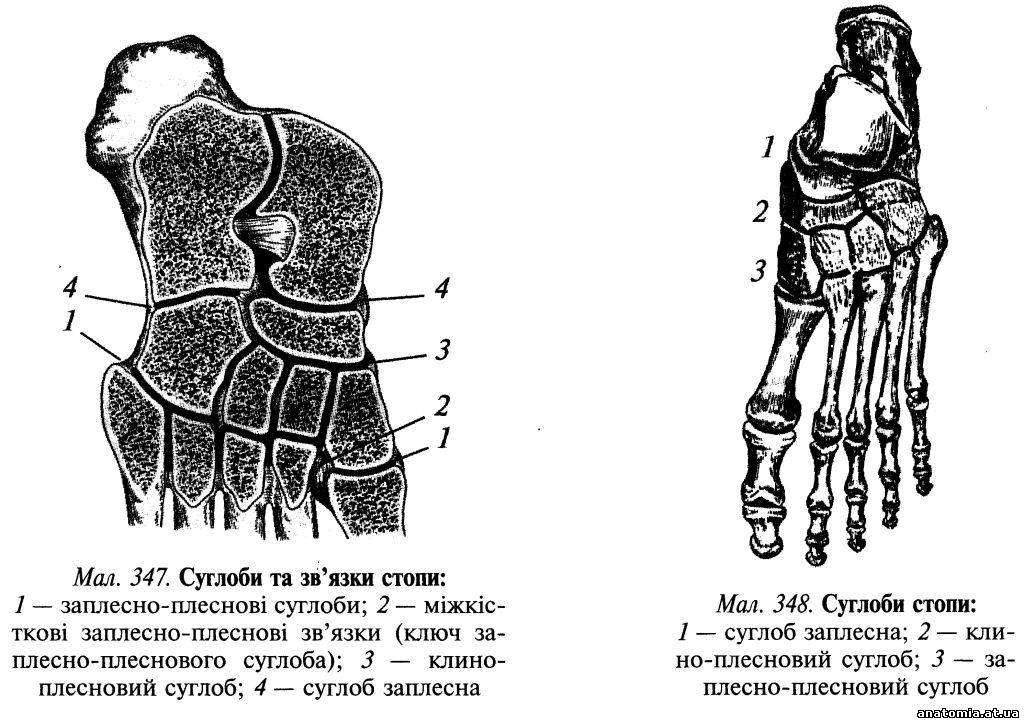 Пошарова топографія. Шкіра підошвової поверхні стопи товста та міцно зрощена з підлеглим підошвовим апоневрозом (aponeurosis plantaris) за допомогою великої кількості сполучнотканинних перегородок, які пронизують підшкірну жирову клітковину. Підшкірна жирова клітковина добре розвинена в ділянці п'яткового горба і головок плеснових кісток, де вона виконує роль амортизатора. Завдяки її вираженій комірковій будові нагнійні проц... Читати далі... |
Розділ: | Категорія: Топографічна анатомія нижньої кінцівки